د کولیسترول بایوسینتیسس او د هغې بایو کیمیکل - ذیابیطس
پرته له شک څخه ، کولیسټرول عام خلکو ته ترټولو غوره پیژندل شوی لیپید دی it دا بدنام دی ځکه چې د وینې د لوړ کولیسټرول او د انسان د زړه ناروغیو تعدد ترمنځ لوړ ارتباط لري. د حجرو غشا د یوې برخې او د سټرایډ هورمونونو او د معدې اسیدونو مخکښ په توګه د کولیسټرول مهم رول ته لږه پاملرنه شوې. کولیسټرول د انسانانو په ګډون د ډیری څارویو لپاره اړین دی ، مګر په تیرو خواړو کې د هغې شتون اختیاري دی - د بدن حجرې پخپله کولی شي دا د ساده مخکښو څخه ترکیب کړي.
د دې 27-کاربون مرکب جوړښت د دې بایوسینتس لپاره یوه پیچلې لاره وړاندیز کوي ، مګر د دې ټول کاربن اتومونه د یو واحد مخکیني لخوا چمتو شوي - اکسیټ. اسوپرین بلاکس - د اکسیټ څخه تر کولیسټرول پورې ترټولو مهم مینځګړیتوب ، دا د ډیری طبیعي لیپایډونو مخکښان دي ، او هغه میکانیزمونه چې د آی اسوپرین بلاکونو له لارې پولیمیریز کیږي په ټولو میټابولیک لارو کې ورته دي.
موږ د اکسیټ څخه د کولیسترول بایوسینتیسس په لاره کې د اصلي مرحلو معاینه کولو سره پیل کوو ، بیا د وینې جریان له لارې د کولیسترول لیږد ، د حجرو لخوا د هغې جذب ، د کولیسټرول ترکیب نورمال تنظیم او د ضعیف جذب یا ټرانسپورټ قضیو کې تنظیم کولو په اړه بحث کوو. بیا موږ نورو موادو ته ګورو چې د کولیسټرول څخه راځي لکه د بائل اسید او سټرایډ هورمونونه. په نهایت کې ، د ډیری مرکبونو رامینځته کولو لپاره د بایوسینتیک لارو بیان - د اسوپرین بلاکونو مشتق ، په کوم کې چې د کولیسټرول ترکیب سره لومړني مرحلې شتون لري ، به په بایوسینتسس کې د اسوپرینایډ کمیدنې غیر معمولي استثنایی توضیح کړي.
کولیسټرول په څلور مرحلو کې د اسټیل CoA څخه تولید کیږي
کولیسټرول ، لکه د اوږدې زنځیر غوړ اسیدونه ، د اسټیل CoA څخه جوړ شوي ، مګر د مجلس ب patternه په بشپړ ډول توپیر لري. په لومړیو تجربو کې ، اسیټټ د 14 سی سره لیبل شوی په میتیل یا کاربوکسیل کاربن اتوم کې د څارویو خواړه ته اضافه شوي. په کولیسټرول کې د لیبل د توزیع پراساس د څارویو دوه ډلو څخه جلا (عکس 21-22) ، د کولیسټرول بایوسینتیسز انزیمیک مرحلې بیان شوې.
عکس 21-32. د کولیسټرول د کاربن اتومونو سرچینه. د راډیو اکټیو اکسیټ کارولو د تجربو پرمهال پیژندل شوی د میتیل کاربن (تور) یا کاربوکسیل کاربن (سور) سره لیبل شوی. په ککړ جوړښت کې ، حلقې د A نه تر D تورو لخوا ښودل کیږي.

ترکیب په څلورو مرحلو کې واقع کیږي ، لکه څنګه چې په عکس کې ښودل شوي. -3 21- (3: (a) د میسیونټیټ شپږ کاربن منځګړیتوب لپاره د دریو اسیتیټ پاتې شونو کمیدنه ، ()) د فعال isoprene بلاکونو ته mevalonate اړول ، ()) د five 30 کاربن isoprene واحدونو پولیمیرزاسیون کول ترڅو د-30 کاربن لاین طبقه بند رامینځته شي ، ()) د طبقې څرخ ته څرخیدنه د سټرایډ نیوکلیو څلور حلقې ، د کولیسټرول رامینځته کیدو سره د یو لړ بدلونونو (آکسیډریشن ، لرې کولو یا مهاتیل ګروپونو مهاجرت) وروسته.
عکس 21-33. د کولیسترول بایوسنتز عمومي عمومي انځور. په ترکیب کې د ترکیب څلور مرحلو بحث شوی. په سکویلین کې د آئوپرین بلاکونه د سور ویجاړ شوي کرښو لخوا نښه شوي.

مرحله (1). د ایزیټ څخه د میالوونټ ترکیب. د کولیسټرول بایوسینتیسس لومړی مرحله د مینځ مینځ محصول رامینځته کیدو لامل کیږي میولوناټ (عکس 21-34). دوه اسیتیل CoA مالیکولونه د Acetoacetyl CoA ورکولو لپاره سنجول کیږي ، کوم چې د دریم اسیتیل CoA مالیکول سره سنګسروي ترڅو د شپږ کاربن مرکب جوړ کړي β-هایډروکسي β-میتیلګلوټریل - CoA (HM G -CoA). دا دوه لومړني عکس العمل کتل شوي دي thiolase او په ترتیب سره د NM G -CoA ترکیب. سیټوسولیک د NM G-CoA ترکیب دا میټابولیک لاره د میتوچونډریل اسوینزیم سره توپیر لري ، کوم چې د کیټون بدنونو رامینځته کولو پرمهال د NM G -CoA ترکیب تیروي (عکس 17-18 وګورئ).
عکس 21-34. د اسیلیل CoA څخه د میالوونایټ جوړښت. د سیټیل CoA څخه د C-1 او C-2 میالوونیت سرچینه په ګلابي کې روښانه شوې.

دریم عکس العمل د ټولې پروسې سرعت محدودوي. پدې کې ، NM G -CoA mevalonate ته کم شوی ، د کوم لپاره چې د N the D دوه PH مالیکولونه دوه برقیان چمتو کوي. د HMG-CoA رډوکټیس - د ارامه ای آر مدغم ممبر پروټین ، دا خدمت کوي ، لکه څنګه چې موږ به وروسته وګورو ، د کولیسټرول جوړښت میټابولیک لارې تنظیم کولو اصلي ټکي په توګه.
مرحله (2). د mevalonate دوه فعال isoprene ته اړول. د کولیسټرول ترکیب په راتلونکي مرحله کې ، د فاسفیت درې ډلې د ATP مالیکولونو څخه mevalonate ته لیږدول کیږي (شکل 21-35). فاسفیت په C-3 میالوونټ کې د هایدروکسیل ګروپ پورې مربوط 3-فاسفو - 5-pyrophosphomevalonate یو ښه پریښودونکی ګروپ دی ، په راتلونکی مرحله کې دا دواړه فاسفټ او د کاربوکسیل ګروپ دواړه رخصت کیږي ، چې په پنځه کاربن محصول کې دوه ګونی جوړیږي ∆ 3 -isopentenyl pyrophosphet. دا د دوه فعال isoprenes لومړی دی - د کولیسترول ترکیب کې اصلي برخه اخیستونکي. د Δ 3 -isopentenylpyrophosphet د isomeriization دویم فعال isoprene ورکوي ډاییمیټالیل پیروفاسفیت. د نبات حجرو سایټوپلسم کې د isopentenyl pyrophosphet ترکیب د دلته بیان شوي لارې سره سم واقع کیږي. په هرصورت ، د کښت کلوروپلستونه او ډیری باکتریا د میالوونیت څخه خپلواکه لاره کاروي. دا بدیل لاره په څارویو کې نه موندل کیږي ، نو دا د نوي انټي بیوټیکونو رامینځته کولو په وخت کې زړه راښکونکي دی.
عکس 21-35. د میالوونټ بدلول په فعاله اسوپریني بلاکونو کې. شپږ فعال شوي واحدونه د سکالوین جوړولو لپاره ترکیب کوي (شکل 21-36 وګورئ). د 3-فاسفو-5-پیرافوفومومولوټینټ پریښودو ډلو په ګلابي کې روښانه شوي. په مربع قوسونو کې فرضي یا منځګړیتوب دی.

مرحله (3). د شپږ فعال isoprene واحدونو راټولول ترڅو اسکیلین رامینځته کړي. اسوپینټیل پایرفوسفیټ او ډییمیتیالیل پیروفاسفیت اوس د سر څخه تر پونډه رګیدو څخه تیریږي ، په کوم کې چې د پیرو فاسفیټ ګروپ حرکت کوي او د 10 کاربن سلسلې ب formsې - geranyl pyrophosphate (عکس 21-36). (پیروفوسفیټ سر ته رسیدلی.) جیرانیل پیرفاسفیت د اسوپینټیل پایرفاسفیت ، او د 15 کاربن منځګړیو ب headو سره د لاندې نه تر پونډې څخه تر سره کیږي. farnesyl pyrophosphate. په نهایت کې ، د فارنیسیل پایرفوسفیټ دوه مالیکولونه “له سر څخه تر سر” سره یوځای کیږي ، دواړه د فاسفیت ګروپونه لرې کیږي - جوړ شوی اسکیلین.
عکس 21-36. د اسکیلین جوړښت. د 30 کاربن اتومونو لرونکی یو سکوالیین جوړښت د isoprene (پنځه کاربن) بلاکونو په واسطه فعال د غلیظ کولو په جریان کې پیښیږي.

د دې مینځګړیو لپاره عام نومونه د سرچینو نومونو څخه راځي چې له کوم ځای څخه دوی لومړی جلا شوي. جیرانول ، د ګلاب غوړ برخه ده ، د جیرانیم بوی لري ، او فارنیسول ، چې د اکاسیه فارنها رنګونو کې موندل شوی ، د وادی بو ب aه لري. ډیری د طبیعي بوټو بوی د اسوپرین بلاکونو څخه جوړ شوي مرکباتو پورې اړه لري. اسکالین ، لومړی د شارک جگر (سکالوس ډوله) څخه جلا شوی ، د 30 کاربن اتومونو څخه جوړ شوی دی: په مین ځنځیر کې 24 اتومات او د فلزاتو زیرمو کې شپږ اتومونه.
مرحله (4). د سټرایډ نیوکلیو په څلورو حلقو کې د سکوالیین بدلون. په انجیر کې. 21-37 دا په ښکاره ډول لیدل کیږي چې د سکایلین سلسلې جوړښت ، او سټرولونه - سایکلیک. ټول سټرایولونه څلور غلیظ حلقې لري چې د سټرایډ نیوکلیو جوړوي ، او دا ټول الکوهولونه دي په C-3 اتوم کې د هایدروکسیل ګروپ سره ، له همدې امله انګلیسي نوم سټیرول دی. د عمل لاندې سکویلین مونو آکسیجن د O څخه یو د اکسیجن اتوم د اسکیلین سلسلې پای کې اضافه کیږي 2 او یو epoxide جوړیږی. دا انزایم بل مخلوط - فعالیت اکسیډیز دی (21-1 اضافه کوي) ، NADPH د O څخه بل آکسیجن اتوم کموي 2 ته2 او. د محصول دوه برابره اسکالین-2،3-epoxide تنظیم شوی ترڅو د پام وړ متناسب عکس العمل د اسکیلین ایپوکسایډ سلسله د چریک جوړښت ته واړوي. د څارویو په حجرو کې ، دا سایکلینګ د رامینځته کیدو لامل کیږي لانوسټرول چې د سټرایډ نیوکلیوس څلور حلقې ب characterه لري. د پایلې په توګه ، لانوسټرول د شاوخوا 20 تعاملاتو لړیو له لارې په کولیسټرول بدل شو ، چې پدې کې د ځینې فلزي ډلو مهاجرت او د نورو لرې کول شامل دي. د بایوسینتیس د دې حیرانونکې لارې توضیحات ، چې د پیژندل شویو ترمنځ یو له خورا ستونزمن څخه دی ، د 1950 په وروستیو کې د کانراډ بلاچ ، تیوډور لینن ، جان کارنفورټ او جورج پاپیاک لخوا جوړ شوی و.
عکس 21-37. د حلقې بندول د خطي کڅوړې کنډز شوي سټرایډ کور ته بدلوي. لومړۍ مرحله د اکسایډیس لخوا مخلوط فنکشن (مونو اکسیژنیز) سره کتل کیږي ، د کوم چې کاسبریټ N AD PH دی. محصول یو epoxide دی ، کوم چې په راتلونکي مرحله کې د سټرایډ کور رامینځته کولو لپاره سایکل کوي. د څارویو حجرو کې د دې تعاملاتو وروستی محصول کولیسټرول دی؛ په نورو ارګانیزمونو کې سټرولونه له هغې څخه لږ توپیر لري.


کولیسټرول د څارویو حجرو ، نباتاتو ، فنګسونو او سټراټیسټونو یو سټراول ځانګړتیا ده چې ورته ورته ورته سټراولونه تولیدوي.
دوی د اسکیلین - 2،3-ایپوکسایډ ته ورته ترکیب لاره کاروي ، مګر بیا لارې یو څه بدلون کوي ، او نور سټیرولونه رامینځته کیږي ، لکه په ډیری نباتاتو کې سیګمسټرول او په فنګس کې اریګوسټرول (عکس 21-27).
د سکیلین ترکیب لپاره د انرژۍ لګښتونه 21-1
د یوه طبقه مالیکول ترکیب لپاره د انرژي لګښتونه (د ATP مالیکولونو په توګه څرګند شوي) کوم دي؟
حل. د اسیلیل CoA څخه د سکویلین په ترکیب کې ، ATP یوازې هغه مرحله کې مصرفیږي کله چې میالوونټیټ په فعاله اسوپرین سکوالیین مخکیني کې بدل شي. د اسکالوین مالیکول رامینځته کولو لپاره شپږ فعال isoprene مالیکولونو ته اړتیا ده ، او د هر فعال مالیکول تولید لپاره درې د ATP مالیکولونه اړین دي. په ټوله کې ، د 18 ATP مالیکولونه د یو اسکیلین مالیکول ترکیب باندې مصرف شوي.
په بدن کې د کولیسترول مرکبونه
په فقراتو کې ، د کولیسټرول لوی مقدار په ځګر کې ترکیب کیږي. هلته د ځینې کولیسټرول ترکیب شوی د هیپټوسایټونو غشا کې ترکیب شوی ، مګر دا په عمده توګه د دې دریو شکلونو څخه یو کې صادر کیږي: بیلیري (د بائل) کولیسټرول ، د معدې اسید یا د کولیسترول اسټرونه. د معدې اسیدونه او د دوی مالګه د کولیسترول هیدروفیلیک مشتق دی ، کوم چې په ځیګر کې ترکیب کیږي او د شحمو هضم کې مرسته کوي (شکل 17-1 وګورئ). د کولیسټرول Esters په عمل کې په ځګر کې رامینځته کیږي اکیل - CoA-Cholesterol-acyltransferase (ACAT). دا انزایم د کاینزایم A څخه د فایټ اسید پاتې شونو لیږد لیري کولیسټرول هایډروکسیل ګروپ ته لیږدوي (شکل 21-38) ، کولیسټرول په هایدروفوبیک شکل بدلوي. په پټ شوي لیپوپروټین ذرو کې د کولیسترول ایسټرونه د کولیسټرول په کارولو سره نورو نسجونو ته لیږدول کیږي یا په ځیګر کې زیرمه کیږي.
عکس 21-38. د کولیسترول ایسټرونو ترکیب. اتفاقی کولیسټرول د ذخیره کولو او ټرانسپورټ لپاره حیدرابوبیک لا نور ب makesه کوي.

کولیسټرول د غشا د ترکیب لپاره د ودې څاروی ارګانیزم ټولو نسجونو لپاره اړین دي ، او ځینې ارګانونه (د بیلګې په توګه ، د ادرینال غدود او جنسي غدود) د سټرایډ هورمونونو د مخکیني په توګه کولیسټرول کاروي (دا به لاندې بحث شي). کولیسټرول د ویټامین ډي مخکینۍ برخه هم ده (شکل 10-10 ، v. 1).
کولیسټرول او نور لیپایډز پلازما لیپوپروټین لري
د کولیسټرول او د کولیسټرول ایسټرونه ، لکه د ټرایسیګلیسرولونو او فاسفولپایډونو په عملي ډول په اوبو کې د حل وړ نه دي ، په هرصورت ، دوی باید له هغه نسج څخه حرکت وکړي چیرې چې دوی نسجونو ته ترکیب شوي وي چیرې چې دوی به زیرمه شي یا مصرف شي. دوی د وینې جریان لخوا د شکل په توګه لیږدول کیږي د وینې پلازما لیپوپروټین - د ځانګړي کیریر پروټینونو macromolecular پیچلتیاوې (اپوليپوپروټینز) د فاسفولپایډونو ، کولیسټرول ، کولیسټرول ایسټرونو او ټرای سیریلګلیروسولونو سره په دې پیچلو کې په بیلابیلو ترکیبونو کې شتون لري.
اپولوپوپروټینز ("اپپو" پخپله پخپله د لیپایډ پروټین ته اشاره کوي) د لپایډونو سره یوځای کوي ترڅو د لیپوپروټین ذرات ډیری برخې رامینځته کړي - په مرکز کې د هایډروفوبیک لیپایډونو سره د ګرمو پیچلو او په سطح کې هایډروفایلیک امینو اسید ځنځیرونه (عکس. 21-39 ، a). د لپایډونو او پروټینونو مختلف ترکیبونو سره ، د مختلف کثافتونو ذرات رامینځته کیږي - له کلسیمرو څخه تر لوړ کثافت لیپوپروټین پورې. دا ذرات د الټراسنټرافیګریشن (جدول 21-1) لخوا جلا کیدلی شي او لیدلوری د الکټرون مایکروسکوپي په کارولو سره لیدل کیږي (شکل 21-39 ، b). د لیپوپروټینونو هره برخه یو ځانګړی دنده ترسره کوي ، کوم چې د ترکیب ، لیپایډ ترکیب او اپولوپروټین مینځپانګې لخوا ټاکل کیږي. لږترلږه 10 مختلف apolipoproteins د انسان د وینې پلازما کې وموندل شول (جدول 21-2) ، چې اندازه یې توپیر لري ، د ځانګړي انټي باډونو سره عکس العملونه ، او د لیپوپروټینونو مختلف کلاسونو کې ځانګړتیا ویش. دا د پروټین برخې اجزا د نښو په توګه عمل کوي چې لیوپروټین ځانګړو نسجونو ته لارښوونه کوي یا انزایمونه فعالوي چې په لیپوپروټینونو عمل کوي.
جدول 21-1. د انسان پلازما لیپوپروټین
ترکیب (ډله ایز کسر ،٪)
r = 513،000). د LDL یوه ذره د کولیسټرول ایسټرونو شاوخوا 1،500 مالیکولونو کور لري ، د کور شاوخوا شاوخوا د کولیسټرول mo 500lec مالیکولونه ، د فاسفولیپیډونو mo 800 mo مالیکولونه او د apoB-100 یو مالیکول لري. b - د لیپوپروټین څلور ټولګی ، د بریښنایی مایکروسکوپ سره څرګندیږي (د منفي څرګندیدو وروسته). کلاک ویز ، د پورتنۍ کی figureې اندازې څخه پیل کیږي: کلسیمیکران - د 50 څخه 200 nm قطر سره ، PL O NP - له 28 څخه تر 70 nm پورې ، HDL - له 8 څخه تر 11 nm پورې ، او LDL - له 20 څخه تر 55 nm پورې. د لیپوپروټین ملکیتونه په میز کې ورکړل شوي. 21-2.

کلسیمرو د. 17 ، د خواړو ټرایسیګلیسرولونه د کولمو څخه نورو نسجونو ته انتقال کړئ. دا ترټولو لوی لیپوپروټین دی ، دوی د ټریسایلګلیسرولونو ترټولو ټیټ کثافت او ترټولو لوړ نسبي مینځپانګه لري (عکس 17-2 وګورئ). Chylomicrons د اپیټیلیل حجرو په ER کې ترکیب شوی چې د کوچني کولمو استر کوي ، بیا د لیمفاټیک سیسټم له لارې حرکت کوي او د کی sub سبکلویون رګ له لارې د وینې جریان ته ننوځي. Chylomicron apolipoproteins apoB-48 (د دې ټولګیو لیپوپروټین لپاره ځانګړی) ، apoE او apoC-II لري (جدول 21-2). AroC-II د ایډپوز نسج ، زړه ، سکلیټي عضلات ، او د رحم تیغ لرونکي غدې په کیپلیري کې لپو پروټین لیپاز فعالوي ، دې نسجونو ته وړیا غوړ اسیدونه چمتو کوي. پدې توګه ، کلسیمومي د خوراکي غوړ اسید نسجونو ته لیږدوي چیرې چې دوی به مصرف شي یا د سونګ توکو په توګه زیرمه شي (عکس 21-40). د کلسیمون باقیات (په عموم ډول د ټرایسیګلیسرول څخه آزاد دي ، مګر لاهم د کولیسټرول ، apoE او apoB-48 لري) د وینې جریان له لارې ځیګر ته لیږدول کیږي. په ځیګر کې ، رسیپټرې د Chylomicron پاتې شونو کې موجود apoE پورې تړلي او د اندوسیټوسیس لخوا د دوی جذب منځګړیتوب کوي. په هیپاټوسایټونو کې ، دا پاتې شونو کولیسټرول خوشې کوي چې دوی پکې لري او په لیزومومونو کې ویجاړ شوي.
جدول 21-2. د انسان پلازما لیپوپروټین اپولیوپروټین
فنکشن (که پیژندل شوی)
L CAT فعالوي ، د ABC لیږدونکي سره اړیکه لري
د L CAT مخنیوی کوي
د L CAT ، د کولیسترول ټرانسپورټ / تصفیه فعالوي
د LDL ریسیپټر پورې تړاو لري
Chylomicrons ، VLDL ، HDL
Chylomicrons ، VLDL ، HDL
Chylomicrons ، VLDL ، HDL
د VLDL او Chylomicron پاتې شونو پاکولو پیل کوي
کله چې خواړه ډیر غوړ اسیدونه ولري دا مهال د سونګ توکو په توګه کارول کیدی شي ، دوی په ځیګر کې ټرایسیګلیسرولونو بدلوي ، کوم چې د ځانګړي اپولوپروټینونو سره یوه برخه جوړوي ډیر ټیټ کثافت لیپوپروټین (VLDL). په ځیګر کې ډیر کاربوهایډریټونه هم په ټرایسیګلیسرولونو بدلیدلی شي او د VLDL په توګه صادر شوي (شکل 21-40 ، a).د ټرای سیلوګلیروسولونو برسیره ، د VLDL فریکشن یو ځانګړی مقدار د کولیسترول او کولیسترول اسټرونه لري ، په بیله بیا apoB-100 ، apoC-1 ، apoC-II ، apoC III او apoE (جدول 21-2). دا لیپوپروټین هم د وینې له لارې د ځيګر څخه عضلاتو او ایډپوز نسج ته لیږدول کیږي ، چیرې چې وروسته د لیپوپروټین لایپیس د آپ-سي II لخوا فعال کیږي ، وړیا غوړ اسیدونه د VLDL کسر له Triacylglycerols څخه خوشې کیږي. اډیپوسایټس وړیا غوړ اسیدونه نیسي ، بیا یې په ټرایسیګلیسرولونو بدلوي ، کوم چې په دې حجرو کې د لیپایډ انکسیژن (څاڅکي) په توګه ساتل کیږي ، مایوسایټس ، برعکس ، فوري اسیدونه د انرژي تولید لپاره فوري اکسیډ کوي. ډیری د VLDL پاتې شونې د هیپاټوټایټس لخوا د دوران څخه له مینځه وړل کیږي. د دوی جذب ، د کلسیمومونو جذب ته ورته ، د رسیپټرو لخوا مینځګړیتوب کیږي او د VLDL پاتې شونو کې د apeE شتون پورې اړه لري (په 21-2-2 کې ، د apeE او الزایمر ناروغۍ تر مینځ اړیکه بیان شوې).
عکس 21-40. لیوپروټین او د لیپایډ ټرانسپورټ ، او - لیپید د وینې جریان لخوا د لیپوپروټینونو په شکل لیږدول کیږي ، کوم چې د مختلف پروتوکونو او پروټینونو او لپایډونو مختلف ترکیبونو (ټب. 21-1 ، 21-2) سره یوځای کیږي او د دې برخو کثافت سره مساوي دي. د خواړو لیپایډونه په کلسیمونو کې راټولیږي ، ډیری ټرایسیسیلګولروسونه چې په دوی کې شامل دي د لیپوپروټین لایپیس لخوا په کیپیلریانو کې د ایډپوز او د غړو عضلاتو ته خوشې کیږي. د کلسیمون باقیات (په عمده ډول پروټین او کولیسترول لري) د هیپټوسایټس لخوا نیول شوی. د ځيګر څخه انډروجنس لپایډونه او کولیسټرول د VLDL په ب adه اډیپوز او د غړو عضلاتو ته استول کیږي. د VLDL څخه د لپایډونو خوشې کول (د ځینې اپولوپروټینونو له لاسه ورکولو سره) په تدریجي ډول VLDLP LDL ته واړوي ، کوم چې کولیسترول له هايډیټیټیک نسجونو ته رسوي یا ځيګر ته بیرته راستنوي. ځیګر د ریسیپټر منځګړیتوب اندوسیټوسیس لخوا د VLDL ، LDL او پاتې پاتې شیمیکروسون پاتې شوي توکي راوباسي. په خارج هايډیټک نسجونو کې ډیر کولیسټرول د LDL په ب theه بیرته جگر ته لیږدول کیږي. په ځیګر کې ، د کولیسترول یوه برخه په صفر مالګو بدلیږي. b - د وینې پلازما نمونې د لوږې وروسته (کی)) او د لوړ غوړ مینځپانګې (ښي خوا) سره خواړه وروسته وروسته اخیستل شوي. د غوړ خواړو په خوړلو سره رامینځته شوي کلیمیکروون شیدو ته پلازما یو بهرنی ورته والی ورکوي.

د Triacylglycerols له لاسه ورکولو سره ، د VLDL یوه برخه VLDL پاتې شونو ته بدلیږي ، چې د مینځنۍ کثافت لیپوپروټینز (VLDL) هم ویل کیږي ، له VLDL څخه د Triacylglycerols لرې کول د ټیټ کثافت لیپوپروټین (LDL) (ټب. 21-1). د LDL کسر ، کوم چې د کولیسترول او کولیسترول په لویه بډایه بډایه دی ، او همدارنګه apoB-100 لري ، کولیسټرول د هایدیټیټیک نسجونو ته لیږدوي چې ځانګړي رسیپټرونه د دوی پلازما جھلی باندې apoB-100 پیژني. دا اخیستونکي د کولیسټرول او کولیسټرول ایسټرونو پورته کولو منځګړیتوب کوي (لکه څنګه چې لاندې تشریح شوي).
اضافه 21-2.د ApoE ایلیلز د الزیمر ناروغۍ پیښې ټاکي
په انساني نفوس کې ، د ژن کوډینګ اپولوپروټین E درې پیژندل شوي تغیرات (درې ایلییلونه) شتون لري. د AUPE له هرډول څخه ، APOEZ ایلیل په انسانانو کې ترټولو عام دی (نږدې 78٪) ، APOE4 او APOE2 ایلیل په ترتیب سره 15 او 7٪ دي. د APOE4 ایلیل په ځانګړي ډول د الزیمر ناروغۍ لرونکي خلکو ځانګړتیا ده ، او دا اړیکې د لوړې احتمال سره د ناروغۍ پیښې وړاندوینې ته اجازه ورکوي. هغه خلک چې APOE4 په میراث پاتې دي د الزایمر ناروغۍ ناوخته وده خطر لري. د APOE4 لپاره همجنګیس خلک د دې ناروغۍ وده 16 ځله ډیر دي ، د ناروغانو اوسط عمر شاوخوا 70 کاله عمر لري. د هغو خلکو لپاره چې د AROEZ دوه کاپي میراث لري ، په برعکس ، د الزایمر ناروغۍ اوسط عمر 90 کاله څخه ډیر وي.
د apoE4 او الزایمر ناروغۍ تر مینځ د اتحادیې لپاره مالیکولي اساس لاهم نامعلوم دی. سربیره پردې ، دا لاهم روښانه نده چې apoE4 څنګه کولی شي د امایلایډ رګونو په ودې اغیزه وکړي ، کوم چې ظاهرا د الزیمر ناروغۍ اصلي لامل دی (شکل 4-31 ، v. 1 وګورئ). انګیرنې د نیورونونو د سایټسوکلیټون جوړښت ثبات کې د اپو ممکنه رول باندې تمرکز کوي. د apoE2 او apoEZ پروټینونه د نیورونونو مایکروټوبیوالونو پورې اړوند یو شمیر پروټینونو پورې تړاو لري ، پداسې حال کې چې apoE4 پابند ندي. دا کولی شي د نیورون مړینه ګړندي کړي. هر هغه څه چې دا میکانیزم ممکن پای ته ورسیږي ، دا مشاهدې د اپولوپروټینونو بیولوژیکي دندو په اړه زموږ د پوهې پراختیا ته امید ورکوي.
د لیپوپروټین څلورم ډول - لوړ کثافت لیپو پروټین (HDL) ، دا کسر په ځيګر او کوچنۍ کولمو کې د کوچني پروټین لرونکي ذراتو په ب formedه رامینځته کیږي چې نسبتا لږ کولیسټرول لري او په بشپړ ډول د کولیسټرول اسټرونو څخه خالي دی (شکل 21-40). د HDL برخه برخه apoA-I ، apoC-I ، apoC-II او نور apolipoproteins لري (جدول 21-2) ، او همدارنګه د لیتین - کولیسټرول - اکیلټرانسیفرایس (LC AT) ، کوم چې د لیسیټین (فاسفیټایډیلولین) او کولیسټرول (عکس 21-21) څخه د کولیسټرول ایسټرونو جوړښت لیري کوي. L CAT د نوي جوړ شوي HDL ذراتو په سطح کې د Chylomicron کولیسټرول او فاسفټایډیلکولین او VLDL پاتې برخې د کولیسټرول ایسټرونو ته واړوي ، کوم چې د نیوکلیوس جوړیدو پیل کوي ، نوي جوړ شوي ډیسک اکسایډ HDL ذراتو ته په کامل Spherical HDL ذراتو بدلوي. دا د کولیسټرول بډایه لیوپروټین بیا جگر ته راستون کیږي ، چیرې چې د کولیسټرول "خارج" کیږي ، د دې ځینې کولیسټرول په صفرا مالګه بدلیږي.
عکس 21-41. هغه تعامل چې د لیسیتین - کولیسترول - اکیلټرانسفیرز (L CAT) لخوا کتل شوی. دا انزایم د HDL ذرو په سطح کې شتون لري او د apoA-1 لخوا فعال شوی (د HDL فریکشن یوه برخه). د کولیسترول اسټرونه د نوي جوړ شوي HDL ذراتو کې دننه کیږي ، دوی په بالغ HDL بدلوي.

HDL کیدی شي د ریسیپټر منځګړیتوب اینڈوسیټوس په واسطه په ځیګر کې جذب شي ، مګر لږترلږه د HDL کولیسټرول یو شمیر نورو میکانیزمونو نورو نسجونو ته سپارل کیږي. د HDL ذرات کولی شي د SR - BI ریسیپټرو پروټینونه د ځيګر حجرو پلازما جھلی او په سټرایډروجنیک نسجونو لکه ادرنال غدود کې وتړي. دا اخذونکي د اندوسیټوسیز منځګړیتوب نه کوي ، مګر په حجره کې د کولیسترول او د HDL فریکشن نورو لیپایډونو جزوي او انتخابي لیږد. د "ختم شوي" HDL برخې بیا بیا د وینې جریان ته ننوځي ، چیرې چې دا د Chylomicrons او VLDL پاتې شونو څخه د لیپایډ نوي برخې شاملوي. ورته HDL کولی شي په خارج کې نسجونو کې زیرمه کولیسټرول ونیسي او له لارې یې ځیګر ته انتقال کړي د کولیسټرول ترانسپورت معکوس کړئ (عکس 21-40). په ریورس ټرانسپورټ ډولونو کې ، د کولیسټرول بډایه حجرو کې د SR-BI ریسیپټرو سره د HDL پایله د تعامل د حجرې له سطحې څخه د کولیسټرول غیر فعال توزیع کوي HDL ذرو ته ، چې بیا کولیسټرول بیرته جگر ته لیږدوي. په بډایه کولیسټرول حجرو کې د ریورس ټرانسپورټ بل ډول کې ، د HDL له مینځه تللو وروسته ، apoA-I د فعال لیږدونکي ، ABC پروټین سره تعامل کوي. ApoA-I (او په احتمالي توګه HDL) د اندوسیټوسیس لخوا جذب شوی ، بیا یې پټ شوی ، د کولیسترول سره ډک شوی ، چې ځيګر ته لیږدول کیږي.
پروټین ABC1 د ډیری درملو وړونکي لوی کورنۍ برخه ده ، دا کیریرونه ځینې وختونه د ABC ټرانسپورټرانو په نوم یادیږي ، ځکه چې دا ټول د ATP - پابند کیسیټونه (ATP - پابند کیسیټونه) لري ، دوی د شپږ ټرانسسیمبران هیلیکسونو سره دوه ټرانسسیمبران ډومینونه هم لري (وګورئ چیپ. .11 ، v. 1). دا پروټینونه په فعال ډول ډیری پلازما ، امینو اسیدونه ، ویټامینونه ، سټرایډ هورمونونه او د پتې مالګې د پلازما جھلي له لارې لیږدوي. د کیریرانو د دې کورنۍ بل استازی د CFTR پروټین دی ، کوم چې د سیسټیک فایبروسس سره زیانمن شوی (وګورئ. د 11-3 ، v. 1.)
د کولیسټرول ایسټرونه د رسیپټر - مینځل شوي اندوسیټوسیس له لارې حجرو ته ننوځي
د وینې جریان کې هر LDL ذره apoB-100 لري ، کوم چې د سطحي ریسیپټرو پروټینونو لخوا پیژندل شوي -د LDL اخیستونکي د حجرو غشا باندې چې د کولیسټرول اخیستلو ته اړتیا لري. د LDL ریسیپټر ته د LDL پابندۍ د اندوسیټوسیس پیل کوي ، له دې امله LDL او د هغې ریسیپټر د اندوزوم دننه حجرو ته حرکت کوي (شکل 21-22). انډوزوم په پای کې د لیزوموم سره فیوز کوي ، کوم چې انزایمونه لري چې د کولیسترول ایسټرو هایډرولائز کوي ، سایټرول کې د کولیسټرول او غوړ اسیدونه خوشې کوي. د LDL څخه ApoB-100 هم د امینو اسیدونو رامینځته کولو لپاره مات شوی چې سیټوسول کې پټ شوی ، مګر د LDL ریسیپټټر د تخریب کیدو څخه مخنیوی کوي او بیرته د LDL اپټایک کې برخه اخیستنې لپاره د حجرې سطحې ته راستون کیږي. ApoB-100 هم په VLDL کې شتون لري ، مګر د دې رسیپټر - پابند ډومین د دې وړتیا نلري چې LDL ریسیپټر ته پابند کړي L LDL ته د VLDLP بدلول د AptB-100 ډومین د ریسیپټر - پابند ډومین رامینځته کوي. دا د وینې د کولیسټرول ترانسپورت لاره او په نښه شوي نسجونو کې د دې ریسیپټر - مینځل شوي اینڈوسیټوسیس د مایکل براون او جوزف گولډسټین لخوا مطالعه شوي.
مایکل براون او جوزف گولډسټین

عکس 21-42. د رسیپټر منځګړیتوب اینڈوسیټوس لخوا د کولیسټرول نیول.

کولیسټرول ، کوم چې پدې توګه حجرو ته ننوځي ، کولی شي په غشا کې یوځای شي یا د ACAT (عکس 21-28) لخوا د سایټسول ذخیره کولو لپاره د لپایډ څاڅکو کې دننه ځای په ځای شي. کله چې د وینې LDL کسر کې کافي کولیسټرول شتون ولري ، نو د اضافي انسټرایسولر کولیسټرول راټول د هغې ترکیب د کچې کمولو سره مخنیوی کوي.
د LDL ریسیپټر هم د AUPE سره تړلی او د ځیګر لخوا د Chylomicrons او VLDL پاتې شونو په پورته کولو کې د پام وړ رول لوبوي. په هرصورت ، که چیرې د LDL ریسیپټرې شتون نلري (د مثال په توګه ، د ورک شوي LDL رسیپټر جین سره په موږک فشار کې) ، د VLDL باقیات او کلسیمیکران لاهم د ځیګر لخوا جذب کیږي ، پداسې حال کې چې LDL جذب نه کیږي. دا د VLDL او د Chylomicron پاتې شونو رسیپټر - منځګړت endocytosis لپاره د مرستې ذخیره سیسټم شتون په ګوته کوي. د ذخیرې اخیستونکو څخه یو د LRP پروټین دی (لیوپروټین ریسیپټر - اړوند پروټین) ، کوم چې د لیپوپروټین ریسیپټرو پورې اړه لري ، کوم چې د apoE او یو شمیر نورو لیګنډونو پورې تړاو لري.
د کولیسترول بایوسینتیس مقرراتو ډیری کچې
د کولیسټرول ترکیب یو پیچلي او د انرژي له پلوه ګرانه پروسه ده ، نو دا روښانه ده چې بدن د کولیسټرول بایوسینتیسس تنظیم کولو میکانیزم لپاره ګټور دی ، کوم چې د خواړو سره راځي د هغه مقدار سربیره بیا ډکوي. په تی لرونکو ژوو کې ، د کولیسټرول تولید د داخلي حجرو غلظت په واسطه تنظیم کیږي
د کولیسټرول او هورمون ګلوکاګون او انسولین. د HMG - CoA ته mevalonate (شکل 21-34) ته د بدلون مرحله د کولیسټرول جوړښت میټابولیک لارې کې سرعت محدودوي (د تنظیم اصلي ټکی). دا عکس العمل د HMG - CoA Redctase لخوا کتل شوی دی. د کولیسټرول کچو کې بدلونونو په ځواب کې مقررات د جین کوډینګ HMG - CoA ریوډیکټیس لپاره د ښکلي لیږد لیږد تنظیمي سیسټم لخوا مینځګړیتوب کیږي. دا جین ، د 20 نورو جینونو کوډینګ انزایمونو سره یوځای چې د کولیسټرول او غیر پروسس شوي غوړ اسیدونو جذب او ترکیب کې دخیل دي د پروټینونو په نوم د کوچني کورنۍ لخوا کنټرول کیږي چې د پروټین جوړیدو سټراول - تنظیمي عنصر سره تعامل کوي (SREBP ، سټیرول تنظیمي عنصر پابند پروټین) . د ترکیب څخه وروسته ، دا پروټین د اینڈوپلازمیک reticula کې معرفي کیږي. یوازینی محلول کیدونکي امینو - ټرمینل SREBP ډومین د Ch په بیان شوي میکانیزمونو کارولو سره د لیږد فعالونکي په توګه دنده ترسره کوي. 28 (v. 3). په هرصورت ، دا ډومین نويکل ته لاسرسی نلري او د جین په فعالیت کې برخه نشي اخیستلی تر هغه چې دا په SREBP ماليکول کې پاتې شي. د HMG جین - CoA ریوډکټیس او نورو جینونو د لیږدونې فعالولو لپاره ، د لیکلې ب activeه فعاله ډومین د پروټولوټیک کفایت په واسطه د SREBP پاتې برخې څخه جلا کیږي. کله چې د کولیسټرول لوړه وي ، د SREBP پروټین غیر فعال وي ، په یوه پیچلي کې د ER سره تثبیت شوي د SCAP په نوم بل پروټین سره (SREBP - د کلیمې فعالولو پروټین) (شکل. 21-43). دا SCAP دی چې کولیسترول او یو شمیر نور سټرایالونه وتړي ، د سټرایټ سینسر په توګه کار کوي. کله چې د سټرول کچه لوړه وي ، د SCAP - SREBP پیچل شاید د یو بل پروټین سره اړیکه ونیسي ، کوم چې په ER کې بشپړ پیچلی ساتي. کله چې په حجره کې د سټراولونو کچه راټیټ شي ، په SCAP کې سازماني بدلون د ساتنې فعالیت له لاسه ورکولو لامل کیږي ، او د SCAP - SREBP پیچلي د ګولۍ پیچلي ته د وازونو دننه لیږدوي. په ګولګي کمپلیکس کې ، د SREBP پروټین دوه ځله د دوه مختلف پروتوزونو پواسطه پاک شوی ، دوهم درواغ د سایټوسول ته د امینو - ترمینل ډومین خوشې کوي. دا ډومین نويکل ته حرکت کوي او د هدف جینونو لیږد فعالوي. د امینو ټرمینل SREBP پروټین ډومین لنډ نیم ژوند لري او په چټکۍ سره د پروټیزوم لخوا تخریب شوی (شکل 27-28 ، t. 3 وګورئ). کله چې د سټرول کچه په کافي اندازه لوړه شي ، د امینو ټرمینز سره د SR EBP پروټین ډومینونو پروتوالیټیک خوشې کول بیا بند شوي ، او د موجوده فعال ډومینونو پروټیزوم تخفیف د هدف جینونو ګړندۍ بندیدو لامل کیږي.
عکس 21-43. د SR EBP فعالیت. د SREB P پروټینونه د سټرایټ تنظیم شوي عنصر (شنه رنګ) سره تعامل کوي ، د ترکیب وروسته سمدلاسه ER ته معرفي کیږي ، د S CAP (سور رنګ) سره یو پیچلي جوړوي. (N او C د پروټینو امین او کاربوکسیل پایونه په نښه کوي.) د S-CAP پابند حالت کې ، د SRE BP پروټین غیر فعال دي. کله چې د سټراول کچه ټیټه شي ، د SR EBP-S CAP پیچلي د ګولګي پیچلې ته کډه کوي ، او د SR EBP پروټینونه په ترتیب سره د دوه مختلف پروټینونو لخوا پاک شوي. د امینو اسید آزاد شوي ترمینل SR EBP پروټین ډومین نويکلیس ته منتقل کیږي ، چیرې چې دا د سټرایټ - تنظیم شوي جینونو لیږد فعالوي.

د کولیسټرول ترکیب د نورو نورو میکانیزمونو لخوا هم تنظیمیږي (شکل 21-44). هورمونل کنټرول د NM G-CoA رډوکټیس د کوالیټ ترمیم لخوا مینځګړیتوب کیږي. دا انزایم په فاسفوریلایټ (غیر فعال) او ډیفاسفوریلاډ (فعال) ب inو کې شتون لري. ګلوګګون د انزایم فاسفریلیشن (غیر فعال کول) هڅوي ، او انسولین ډیفاسفریلیشن هڅوي ، انزایم فعالوي او د کولیسترول ترکیب ملاتړ کوي. د کولیسټرول عالي انسټا سیلولر غلظت ASAT فعالوي ، کوم چې د تویولو لپاره د کولیسټرول تداوی زیاتوي. په نهایت کې ، د سیلولر کولیسټرول لوړې کچې د جین لیږد مخه نیسي چې د LDL ریسیپټر ته داخلیږي ، د دې ریسیپټر تولید کموي او له همدې امله ، له وینې څخه د کولیسټرول لوړول.
عکس 21-44. د کولیسټرول کچه تنظیم کول له خواړو څخه د ترکیب او جذب تر مینځ توازن چمتو کوي. ګلوګګون د NM G -CoA ریډیکټیس فاسفریلیشن (غیر فعال کول) اسانوي ، انسولین ډیفوسفریلیشن (فعالیت) هڅوي. ایکس - ناپیژندل شوی کولیسټرول میتابولیتونه چې د NM G -CoA ریډیکټیس پروټینولیز هڅوي.

غیر منظم کولیسټرول کولی شي په انسانانو کې د جدي ناروغۍ لامل شي. کله چې د خواړو څخه ترلاسه شوي ترکیب شوي کولیسټرول او د کولیسټرول مجموعه د غشا مجلس لپاره اړین مقدار څخه زیات شي ، د وینې رګونو کې سټرویډونو ترکیب ، د وینې په رګونو کې (د ایتروسکلروټیک پلاکونو) رامینځته کیدی شي چې د دوی د بندیدو لامل کیږي (ایتروسکلروسیس). په صنعتي هیوادونو کې ، د زړه د شریانونو د خنډ له امله د زړه ضعیف دی چې د مړینې اصلي لامل دی. د ایتروسکلروسیس وده د وینې کولیسټرول لوړې کچې پورې اړه لري او په ځانګړي ډول د لوړ کولیسټرول سره چې د LDL فرکشن لخوا زغمل کیږي؛ د وینې HDL لوړه کچه ، برعکس ، په مناسب ډول د وینې رګونو حالت اغیزه کوي.
د هایټرو کولیسټرولیمیا (جنیټیکي عیب) سره ، د وینې د کولیسټرول کچه خورا لوړه ده - شدید اییتروسکلروسیس په خلکو کې دمخه په ماشومتوب کې وده کوي. د ناقص LDL ریسیپټر له امله ، د LDL کولیسټرول ریپټر منځګړیتوب نشتون پیښیږي. د پایلې په توګه ، کولیسټرول د وینې جریان څخه نه ایستل کیږي ، دا د ایتروسکلروټیک پلاکونو رامینځته کولو کې برخه اخلي او مرسته کوي. د Endogenous کولیسټرول ترکیب دوام لري ، په وینه کې د اضافي کولیسټرول سربیره ، ځکه چې د حجرو څخه بهر کولیسټرول نشي کولی د حجرو ته ننوځي چې د رګ د لارې ترکیب تنظیم کړي (شکل 21-44).د وینی هایپرکولسټرولیمیا او نورو ناروغیو ناروغانو درملنې لپاره چې د لوړ سیرم کولیسټرول سره تړاو لري ، سټیټین ټولګی کارول کیږي. ځینې یې د طبیعي سرچینو څخه ترلاسه شوي ، پداسې حال کې چې نور یې د درمل جوړولو صنعت لخوا ترکیب شوي. سټیټونه میالوونټ ته ورته دي (21-3 اضافه کړئ) او د NMS-CoA رډوکسټیس سیالي مخنیوی کونکي دي.
ضمیمه 21-3. درملنه. د شحمو مفروضه او د سټیټونو رامینځته کول
کورونري د زړه ناروغي (CHD) په پرمختللو هیوادونو کې د مړینې اصلي لامل دی. د زړه د شریانونو تنګول چې زړه ته وینه لیږدوي د غوړ زیرمو رامینځته کیدو په پایله کې رامینځته کیږي چې د ایتروسکلروټیک پلاکونو په نوم یادیږي these دا تختې د کولیسترول ، فایبرر پروټینونه ، کلسیم ، پلیټلیټ کلاټس او د حجرو ټوټې لري. په XX پیړۍ کې. د شریان خنډ (اتیرسکلروسیس) او د وینې کولیسټرول ترمینځ د اړیکو په اړه فعال بحث شتون درلود. پدې بحثونو او فعالې څیړنې د اغیزمن درملو رامینځته کولو لامل شوي چې کولیسټرول ټیټ کړي.
په 1913 کې ، د روسی پوهانو او تجرباتي رنځپوهنې په برخه کې متخصص ، این. اینیککوف ، یو کار خپور کړ چې پکې هغه په ګوته کوي چې د کولیسټرول بډایه خواړو سره خرگو شوي د وینې رګونو ته زیان رسوي چې د زړو خلکو په رګونو کې د ایتروسکلروټیک پلاکونو سره ورته وي. آنیچکوف خپله څیړنه د څو لسیزو لپاره ترسره کړه او پایلې یې په لویدیځ مشهور پیژندل شوي ژورنالونو کې خپرې کړې. بدبختانه ، د هغه ارقام په انسانانو کې د ایتروسکلروسیس د ودې لپاره اساس نه دی رامینځته کړی ، ځکه چې پدې وخت کې فرضیه موجوده وه چې دا ناروغي د عمر طبیعي پایله ده او مخنیوی یې نشي کیدی. په هرصورت ، شواهد ورو ورو د سیرم کولیسټرول او د ایتروسکلروسیس (لیپید فرضيه) او په 1960 مو کلونو کې د اړیکو تر مینځ اړیکې راټولې کړې وې. ځینې څیړونکو په څرګنده وویل چې دا ناروغي د درملو سره درملنه کیدی شي. په هرصورت ، برعکس لیدلوری په 1984 کې د متحده ایالاتو د روغتیا ملي انسټیټیوټ (د کورونري لومړني مخنیوي محاکمه) لخوا ترسره شوي د کولیسټرول رول پراخه مطالعې پایله پورې خپرولو پورې شتون درلود. د مایکارډیال انفکشن په فریکونسۍ کې احصایوي د پام وړ کمښت او د وینې د کولیسټرول کمیدو سره سټروک ښودل شوي. پدې څیړنه کې ، کولیسټرول ، د anion د تبادلې رال چې د بایټ اسید تړلی ، د کولیسټرول ټیټولو لپاره کارول شوی. پایلې د نوي ، خورا پیاوړي معالجوي درملو لټون هڅوي. زه باید ووایم چې په ساینسي نړۍ کې ، د لیپډ فرضيه د اعتبار په اړه شکونه په بشپړ ډول یوازې د 1980 مو کلونو په وروستیو کې د سټیټینونو په راتګ سره له لاسه ورکړل - د 1990 مو کلونو په لومړیو کې.
لومړی مجسم د اکیرا اندو لخوا په ټوکیو کې په سانکیو کې وموندل شو. اینڈو خپل کار په 1976 کې خپور کړ ، که څه هم هغه د څو کلونو لپاره د کولیسترول میتابولیزم ستونزه حل کړه. په 1971 کې ، هغه وړاندیز وکړ چې د کولیسترول ترکیب مخنیوی کونکي ممکن د هغه وخت مطالعه شوي انټي بیوتیکونو په مشروم تولیدونکو کې هم ولري. د څو کلونو متشبث کار لپاره ، هغه د مختلف مشرومونو 6،000 څخه ډیر کلتورونه تحلیل کړل ، تر هغه چې مثبتې پایلې ته ورسي. پایله کې مرکب کمپیکٹین نومیده. دې مادې په سپیو او بندرونو کې کولیسټرول ټیټ کړی. دې مطالعو د ټیکساس پوهنتون د سویل لویدیز طبي ښوونځي د مایکل براون او جوزف گولډسټین پام ځانته اړولی. براون او گولډسټین د اندو سره یوځای ګډه مطالعه پیل کړه او د هغه ارقام یې تایید کړل. د لومړي کلینیکي آزموینو لوی بریالیتوبونه د دې نوي درملو پرمختګ کې د درملو شرکتونو برخه اخیستې. په میرک کې ، د الفریډ البرټس او روی واګلوس په مشرۍ یوې ډلې د مشروم کلتورونو نوې سکرینینګ پیل کړ او د ټولټال 18 کلتورونو تحلیل کولو په نتیجه کې یو بل فعال درمل کشف کړ. نوې ماده لوواستاتین نومیږي. په هرصورت ، په ورته وخت کې ، دا په پراخه کچه باور درلود چې سپيانو ته د کمپیکٹین د لوړې اندازې اداره کول د سرطان پرمختګ او 1980 کې د نوي مجسمې لټون ته لار هواروي. ځنډول شوی په هرصورت ، پدې وخت کې ، د فیمیل هایپرکولیسټرولیا ناروغانو درملنې لپاره د سټیټینز کارولو ګټې دمخه څرګند وې. د نړیوالو کارپوهانو او د خوړو او درملو اداره (FDA ، USA) سره د ډیری مشورو وروسته ، میرک د لویسټاسټین وده پیل کړه. په راتلونکو دوه لسیزو کې پراخه مطالعې د لوواستاټین کارسنجینیک اثر او د مخدره توکو نوی نسل نه دی څرګند کړی چې وروسته له هغې څرګند شو.
عکس 1. سټیټینز د NM G-CoA رډوکټیس مخنیوی کوي. د میالوونایټ او څلور درملو تولیداتو (سټیټینز) جوړښت پرتله کول چې د NM G -CoA ریډیکټیس فعالیت مخنیوی کوي.

اسټینز د HMG - CoA - Redctase د عمل مخه ونیسئ ، د mevalonate جوړښت مشابه کول ، او پدې توګه د کولیسټرول ترکیب بندوي. د LDL رسیپټر جین یوه کاپي کې د نیمګړتیا له امله د هایپرکولیسټرولیمیا ناروغانو کې ، کله چې لوسټاسټین اخلي ، د کولیسترول کچه 30 reduced کمه شوې. مخدره مواد د ځانګړي رالونو سره په ترکیب کې خورا اغیزناک دي چې د معدې اسیدونه تړلي او د کولمو څخه د دوی بیرته جذب ته مخه کوي.
اوس مهال ، سټیټینز ډیری وختونه د وینې پلازما کولیسټرول ټیټولو لپاره کارول کیږي. کله چې کوم درمل وخورئ ، نو پوښتنه د دوی د نه منلو وړ اړخیزو اغیزو په اړه راپورته کیږي. په هرصورت ، د سټیټونو په قضیه کې ، ډیری اړخیزې اغیزې ، برعکس ، مثبت دي. دا درمل کولی شي د وینې جریان تحرک کړي ، دمخه موجود ایتروسکلروټیک پلاکونه تنظیم کړي (نو دا چې دوی د وینې رګونو دیوالونو څخه نه وځي او د وینې جریان سره مداخله نه کوي) ، د پلیټلیټ ادغام مخنیوی کوي ، او همدارنګه د وینې رګونو دیوالونو کې سوزیدونکي پروسې ضعیفوي. په ناروغانو کې چې د لومړي ځل لپاره سټیټینونه اخلي ، دا اغیزې حتی د کولیسټرول کچه راټیټیدو دمخه څرګندیږي ، او احتمال لري د آیسوپرینایډ ترکیب مخنیوی سره تړاو ولري. البته ، د سټیټینز هر اړخ اغیزې ګټورې ندي. په ځینو ناروغانو کې (معمولا د نورو درملو سره ترکیب کولو کې سټیټین اخیستل چې د کولیسټرول ټیټ دي) ، د غړو درد او د غړو کمزوری کیدی شي ، او ځینې وختونه په کلکه قوي شکل کې وي. د سټیټینز نور خورا ډیر شمیر ضمني اغیزې هم راجسټر شوي ، کوم چې ، له نېکه مرغه ، په ندرت سره پیښیږي. د ناروغانو په لوی اکثریت کې ، د سټیټین اخیستل کولی شي د زړه ناروغۍ پراختیا مخه ونیسي. د نورو درملو په څیر ، سټینز باید یوازې ستاسو د ډاکټر لخوا وړاندیز شوي وکارول شي.

د HDL کولیسټرول د میراث نشتون سره ، د کولیسټرول کچه خورا ټیټه ده ، د ټینګیر ناروغۍ سره ، کولیسټرول په عملي ډول نه ټاکل کیږي. دواړه جینیاتي اختلالات په ABC1 پروټین کې د بدلون له امله رامینځته کیږي. د HDL-free کولیسټرول کسر کولی شي د ABC1 کمبود حجرو څخه کولیسترول ونه نیسي ، او د کولیسترول ضایع شوي حجرې ژر تر ژره له وینې لرې او له مینځه وړل کیږي. د HDL او ټانګیر ناروغۍ دواړه میراثي نشتون خورا ډیر نادر دي (د تانګیر ناروغۍ سره د 100 څخه ډیرې کورنۍ په نړۍ کې پیژندل شوي) ، مګر دا ناروغۍ د HDL پلازما کچه تنظیم کولو کې د ABC1 پروټین رول څرګندوي. څنګه چې ټیټ پلازما د HDL کچه د کورونري شریان زیان سره د لوړې کچې سره ارتباط لري ، د ABC1 پروټین ممکن د HDL کچه تنظیم کولو لپاره ډیزاین شوي درملو لپاره ګټور هدف وي. ■
سټرایډ هورمونونه د کولیسټرول د اړخ زنځیر او د دې اکسیډریشن تقسیم کولو سره رامینځته کیږي.
یو څوک خپل ټول سټرایډ هورمونونه د کولیسټرول څخه لاسته راوړي (عکس 21-45). د سټرایډ هورمون دوه ټولګي په ادرینال کورتکس کې ترکیب شوي: معدني کارټيکوئډز ،کوم چې غیر غیر عضوي ایونونو جذب تنظیموي (Na +، C l - او HC O 3 -) په پښتورګو کې ، او ګلوکوکورټیکوایډز ، کوم چې د ګلوکوزواژینسیس په تنظیم کې مرسته کوي او د انفلاسیون غبرګون کموي. جنسي هورمونونه د نارینه او ښځینه تناسلي حجرو کې او په نبض کې رامینځته کیږي. د دوی په منځ کې پروجسترون چې د ښځينه تناسلي عضلې تنظيموي ، androgens (د مثال په توګه ټیسټورسټون) او ایسټروجنونه (ایسټراډیول) ، کوم چې په ترتیب سره په نارینه او ښځینه کې د ثانوي جنسي ځانګړتیاو پراختیا باندې تاثیر کوي. سټرایډ هورمونونه په خورا ټیټ غلظت کې تاثیر لري او له همدې امله په نسبتا لږ مقدار کې ترکیب کیږي. د بیت مالګو په پرتله ، نسبتا لږ کولیسټرول د سټرایډ هورمونونو تولید لپاره مصرف کیږي.

عکس 21-45. ځینې سټرایډ هورمونونه د کولیسټرول څخه رامینځته کیږي. د دې ځینې مرکب جوړښتونه په عکس کې ښودل شوي. 10-19 ، v. 1.

د سټرایډ هورمون ترکیب د کولیسټرول C-17 D - حلقه "اړخ چین" کې د څو یا ټولو کاربن اتومونو لرې کولو ته اړتیا لري. د غاړې چینې لرې کول د سټرایډروجینیک نسجونو مایټوکونډریا کې واقع کیږي. د حذف کولو پروسه د دوه زنځیر کاربن اتومونو هایډرو آکسیلیشن لري د غاړې سلسلې (C-20 او C-22) ، بیا د دوی تر مینځ د اړیکو پاکیدل (شکل 21-46). د مختلف هورمونونو جوړښت کې د اکسیجن اتومونو معرفي کول هم شامل دي. د سټرایډ بایوسینتس په جریان کې ټول هایډروکسیلریشن او آکسیډریشن تعاملات د مخلوط - فعالیت اکسایډز (21-1 اضافه کول) لخوا کتل کیږي کوم چې NА D PH ، O کاروي 2 او د مایټوکونډرال سایټروکوم P-450.
عکس 21-46. د سټرایډ هورمونونو ترکیب کې د غاړې چین پاکوالي. پدې اکسیدیس سیسټم کې د مخلوط فنکشن سره چې کاربن اټوم ته نږدې اکسیډیز کوي ، سایټروکوم P-450 د بریښنایی کیریر په توګه عمل کوي. همدارنګه په پروسه کې دخیل د بریښنایی ترانسپورت پروټینونه ، اډرینوډوکسین او اډرینوډوکسین رډوکټیس دي. د اړخ زنځیرونو ویشلو دا سیسټم د اډرینال کورټیکس مایټوکونډریا کې وموندل شو ، چیرې چې د سټرایډونو فعال تولید پکې کیږي. Pregnenolone د نورو ټولو سټرایډ هورمونونو مخکښ دی (شکل 21-45).

د کولیسترول بایوسینتیسز مینځګړیتوبونه په ډیری نورو میتابولیک لارو کې دخیل دي.
د کولیسټرول بایوسینتس د مینځمهالي په توګه د دې د رول سربیره ، isopentenyl pyrophosphet د ډیری ژور بایومیوکولونو ترکیب کې د فعال مخکښ په توګه دنده ترسره کوي چې بیلابیل بیولوژیک افعال ترسره کوي (شکل 21-47). پدې کې ویټامینونه A ، E او K ، د نباتاتو رنګونه لکه کیروټین او د کلوروفیل فایټول چین ، طبیعي ربر ، ډیری اړین غوړ (د مثال په توګه ، د نیبو غوړ خوشبويي ، یوکلیپټس ، کښتیو) ، د حشراتو کوچني هارمون چې میټامورفیسس تنظیموي ، ډولوچولز ، کوم چې د پولیساکریډیس ، یوبیوکینون او پلاسټوکینون پیچلي ترکیب کې د لپید - محلول وړونکي کیریر په توګه خدمت کوي - په مایټوکونډریا او کلوروپلاستونو کې د الیکترون کیریر. دا ټول مالیکولونه په جوړښت کې isoprenoids دي. په طبیعت کې له 20،000 څخه ډیر مختلف isoprenoids موندل شوي ، او په سلهاو نوي نوي راپور شوي.
عکس 21-47. د isoprenoids د بایو سینسیټز عمومي انځور. دلته د وړاندې شوي ډیری پای محصولاتو جوړښتونه په فصل کې ورکړل شوي. 10 (v. 1).

Prenylation (د isoprenoid covalent ضمیمه ، شکل 27-25 وګورئ) یو عام میکانیزم دی چې له مخې پروټینونه د تی لرونکي حجرو داخلي سطح باندې لنگر کوي (شکل 11-14 وګورئ). په ځینې پروټینونو کې ، محدود لیپید د 15 کاربن فارنیسیل ګروپ لخوا استازیتوب کیږي ، په نورو کې دا د 20 کاربن ګیرانیل جیرانیل ګروپ دی. دا دوه ډوله لیپایډونه مختلف انزایمونه لري. احتمال شتون لري چې د تیروتنې پروسې پروټینونه مختلف جھلی ته مستقیم پروټینونه په دې پورې اړه لري چې کوم لپید سره تړاو لري. د پروټین پاکول د isoprene ډیرویوټیو لپاره بل مهم رول لري - د کولیسترول میتابولیک لارې برخه اخیستونکي.
د 21.4 برخې لنډیز د کولیسټرول ، سټرایډز ، او اسوپروینایډونو بایوسینتیسس
■ کولیسټرول د منځګړو له لارې د پیچلي تعامل په ترتیب کې د اسټیل - CoA څخه رامینځته کیږي لکه د hydro-هایدروکسای β-میتیلګلوټریل - CoA ، میالوونټ ، دوه فعال isoprene dimethylallyl pyrophosphet او isopentenyl pyrophosphet. د اسوپرین واحدونو ککړتیا غیر سایکلیک سکیلین ورکوي ، کوم چې د رنګه سیسټم او سټرایډ اړخ سلسله رامینځته کولو لپاره سائیکل کوي.
ch د کولیسټرول ترکیب د هورمونل کنټرول لاندې دی او سربیره پردې ، د انسټرایسولر کولیسټرول د زیاتیدونکي غلظت لخوا مخنیوی کیږي ، کوم چې د متقابل تعدیل او د لیږد تنظیم کولو له لارې رامینځته کیږي.
oles کولیسټرول او د کولیسترول اسټرونه د وینې لخوا د پلازما لیپوپروټین په توګه لیږدول کیږي. د VLDL فریکشن کولیسټرول ، د کولیسترول اسټرونه او ټریاسیلګلیروسولونه له ځیګر څخه نورو نسجونو ته لیږدوي ، چیرې چې د ټرایسیګلیسرولونه د لیپو پروټین لیپیس پواسطه پاک شوي وي او VLDL LDL ته اړول کیږي. د کولیسټرول او کولیسټرول ایسټرونو بډایه شوي د LDL کسر په غیر مستقیم ډول د رسیپټرو لخوا د Endocytosis لخوا نیول کیږي ، پداسې حال کې چې په LDL کې B-100 اپالیپوپروټین د پلازما جھلی ریسیپټرو لخوا پیژندل شوی. HDL کولیسټرول له وینې څخه لرې کوي ، جګر ته یې لیږدوي. تغذیه شرایط یا په کولیسټرول میتابولیزم کې جنیټکي نیمګړتیاوې د ایتروسکلروسیز او مایکارډیال انفکشن لامل کیدی شي.
■ سټرایډ هورمونونه (ګلوکوکورټیکوایډز ، مینرالکورټیکوایډز او جنسي هورمونونه) د کولیسټرول څخه د غاړې سلسلې بدلولو او د حلقو سټرایډ سیسټم ته د اکسیجن اتومونو معرفي کولو څخه رامینځته کیږي. ډیری نور اسوپرینوډ مرکبات د کولیسټرول سره د isopentenyl pyrophosphet او dimethylallyl pyrophosphet د کمولو له لارې له mevanonate څخه تولیدیږي.
certain د ځینې پروټینونو ساده کول دوی ته د حجرو غشاګانو سره د تړلو سایټونو لارښوونه کوي او د دوی بیولوژیکي فعالیت لپاره مهم دی.
پوښتنه 48. د لوړ غوړ اسیدونو میتابولیزم تنظیم کول (ox-اکسیډریشن او بایوسینتیس). د مالونیل CoA ترکیب. د اسټیل CoA کاربوکسیلاز ، د دې د فعالیت تنظیم کول. د مایټوکونډریا د داخلي غشا له لارې د اکیل Co-a لیږدول.
اصلي
د فینیلالینین مقدار مصرف شوی
په 2 لارو کې:
بلوي
په مرغیو کې ،
بدلوي
په ټایرسن کې.
بدلول
فینیلایلینین ته په لومړي ډول
د ډیر د لرې کولو لپاره اړین دي
فینیلاالینین ، د لوړ تمرکز راهیسې
دا حجرو ته زهرجن دی. تعلیم
ټیروسین واقعیا اهمیت نلري
ځکه چې د دې امینو اسید کمښت
په حجرو کې په عملي ډول نه پیښیږي.
اصلي
فینیلالانین میتابولیزم پیل کیږي
د دې هایډروآکسیشن سره (شکل 9-29) ، په
د ټایروسین پایله
دا عکس العمل د ځانګړي لخوا کتل کیږي
monooxy-nase - فینیلاالینین هیدرا (ژیسیلاس ،
کوم چې د شریک تولید کونکي په توګه کار کوي
ټیټرایډروبایپټرین (N4BP).
د انزایم فعالیت هم پورې اړه لري
د Fe2 شتون.
په
ځيګر په عمده ډول د خوځښت تحرک دی
ګلایکوژن (برخه 7 وګورئ). په هرصورت
په ځيګر کې ګلایکوجن ختم شوی دی
18-24 ساعته روژه. اصلي سرچینه
ګلوکوز لکه څنګه چې سټاک ختم شوي
ګلیکوګین ګلوکوزجوزنیز کیږي ،
کوم چې د ګړندي کیدو له لارې پیل کیږي
عکس
11-29. لوی میټابولیک بدلونونه
انرژي کله چې جاذب بدل کړي
پوسټ ساسبرټ ایالت. سي ټي
- کیټون باډیز ، FA - غوړ اسیدونه.
4-6 h
وروستی خواړه وروسته سبسټریټس
ګلیسرول د ګلوکوز ترکیب لپاره کارول کیږي ،
امينو اسيدونه او شحمي لکۍ په لوړه کچه
د ګلوکاګون غلظت ترکیب کچه
د غوړو اسیدونو له امله کم شوی
فاسفوریسیون او غیر فعالول
اسټیل CoA کاربوآکسیلاز او نرخ
د پی آکسایډریشن وده کوي. په هرصورت ،
ځيګر ته د غوړ عرضه ډیروي
اسیدونه چې لیږدول کیږي
د غوړ ډیپو څخه اسټیل - CoA رامینځته شوی
د غوړ اسیدونو اکسیدریشن کې ، دا کارول کیږي
په ځیګر کې د کیټون بدنونو ترکیب لپاره.
په
د زیاتوالي غلظت سره د ایډپوز نسج
د ګلوکاګون کم ترکیب اندازه
TAG او لیپولیس تحلیل کیږي. هڅونه
لیپولیس - د فعالولو پایله
هورمون حساس TAG لپیس
د ګلوګګان تر تاثیر لاندې اډیپوسایټونه.
شحمي اسیدونه مهم کیږي
په ځيګر ، غړو او غړو کې د انرژي سرچینې
د زړو نسج
نو
په دې توګه ، د پوسټبرګریشن دوره کې
د وینې ګلوکوز غلظت ساتل کیږي
د 80-100 ملی ګرامه / dl کچه ، او د غوړ په کچه کې
اسیدونه او کیټون بدنونه ډیریږي.
بوره
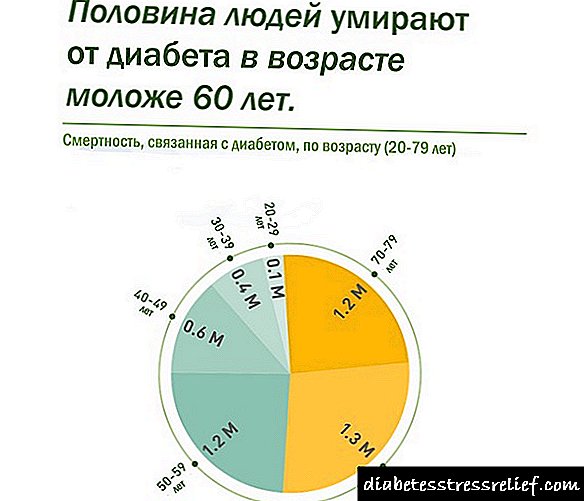
ډايبېتېز يوه ناروغي ده چې پېښېږي
د مطلق یا خپلوۍ له امله
د انسولین کمښت.
الف.
د بورې اصلي کلینیکي ب formsې
د شکرو
د
نړیوال سازمان
د
د توپیرونو پر اساس طبقه بندي
جینیاتی عوامل او کلینیکي
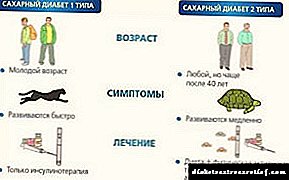
دوه عمده ب :ې: شوگر
د I ډول - د انسولین پورې تړاو لرونکی (IDDM) ، او ډایبېټس
دوهم ډول - غیر انسولین خپلواک (NIDDM).
مقررات
د zhk ترکیب. تنظیمي انزایم
د lcd ترکیب - Acetyl CoA کاربوکسیلاز.
دا انزایم د څو لخوا تنظیمیږي
لارې.
فعالیت / بې لارې کیدل
د انزایم سبونیت کمپلکس. په
د اسټیل CoA کاربوکسلایز غیر فعال ډول
بیلابیل کمپلیکسونه وړاندې کوي ،
چې هر یو یې 4 subunits مشتمل دی.
د انزایم فعالونکی سایټریټ دی. دا هڅوي
د پیچلو ترکیب ، د پایلې په توګه
په دې سره د انزایم فعالیت زیاتیږي
. Inhibitor-palmitoyl-CoA. هغه زنګ ووهي
پيچلي جلا کیدل او کمیدل
د انزایم فعالیت.
فاسفریلیشن / ډیفاسفریلیشن
اسټیل CoA کاربوکسلایز. په
د پوسټ سراسري حالت یا په کې
فزیکي کار ګلوکاګانیز شوی
اډرینالین د اډینالیټ سایکلز له لارې
سیسټم د پروکنیز A او لخوا فعال شوی دی
د سبونیت فاسفوریسیون هڅونه
اسټیل CoA کاربوکسلایز. فاسفوریال
انزایم غیر فعال او د غوړ ترکیب دی
اسيدونه ودريږي
جذب کونکی
د مودې انسولین فاسفیتز فعالوي ،
او اسیتیل - CoA کاربوکسیلاز ته ځي
ډیفیسفوریلیټ شوی حالت. بیا
د سایټریټ اغیز لاندې واقع کیږي
د انزایم د پروټومرو پولیټریک کول ، او
هغه فعاله کیږي. د فعالولو سربیره
انزایم ، سایټریټ یو بل ترسره کوي
د LCD په ترکیب کې فعالیت. جذب کونکی
د ځيګر د حجرو mitochondria موده
سیټریټ راټولوي ، په کوم کې
د اکیل پاتې پاتی کیدو ته لیږدول کیږي
سایتوسول.
مقررات
ox - د آکسیکسیون نرخ.
ox - د آکسیډریشن میتابولیک لاره ،
په کلکه د CPE او عمومي کار سره تړاو لري
د کاتابولیزم لارې. له همدې امله د دې سرعت
د حجرې اړتیا لخوا تنظیم شوی
انرژي i.e. د ATP / ADP او NADH / NAD د تناسب په واسطه ، او همدارنګه د CPE او عکس العمل کچه
د کاتابولیزم عمومي لاره. سرعت
په نسجونو کې اکسیډریشن په موجودیت پورې اړه لري
سبسټریټ ، یعنی.
د غوړ مقدار باندې
اسیدونه میتو کونډریا ته ننوځي.
د وړیا فایټ اسید غلظت
په وینه کې د فعالیدو پرمهال راپورته کیږي
د روژی په جریان کې د ایډپوز نسج کې لیپولیس
د ګلوکاګون تر اغیزې لاندې او د فزیکي پرمهال
د اډرینالین تر اغیز لاندې کار کوي. پدې کې
غوړ اسیدونه کیږی
د بریښنا اصلي سرچینه
د عضلاتو او ځیګر لپاره ، د پایلې په توګه
ox - اکسیډریشن د NADH او اسیتیل CoA مخنیوي لخوا رامینځته کیږي
pyruvate د هایدروجنیز پیچلي.
د pyruvate تشکیل جوړول
له ګلوکوز څخه اسیتیل CoA ته سست دی.
منځګړیتوب میتابولیز راټولیږي
ګلاکولیزس او په ځانګړي ډول ګلوکوز -6-فاسفیت.
ګلوکوز -6-فاسفیت د هیکوسیناس مخه نیسي
او له همدې امله مخنیوی کوي
په پروسه کې د ګلوکوز کارول
ګلیکولیس. له همدې امله ، مخکښ
د اصلي سرچینې په توګه د LCD کارول
د عضلاتو نسج او ځيګر کې انرژي
د عصبي نسجونو لپاره ګلوکوز وژغوري او
د وینې سره حجرې
د اکسیډریشن کچه هم
د انزایم فعالیت پورې اړه لري
کارنیټین اکیلټرانسفرایسز I.
په ځیګر کې ، دا انزایم مخنیوی کیږي.
ملونيیل CoA ، یو ماده جوړه شوې
د lcd بیوسینتیسس سره. د جاذبې په دوره کې
ګلاکولیسس په ځیګر کې فعال او فعال دی
د اسیتیل - CoA تشکیل ډیریږي
د pyruvate څخه. لومړی ترکیب تعامل
د ایسیلیل CoA ته LLD د مالونیل CoA ته اړول.
مالونیل - CoA د Lcd اکسایډریشن مخه نیسي ،
کوم چې د ترکیب لپاره کارول کیدی شي
غوړ.
تعلیم
د ایسټیل CoA - مقرراتو څخه مالونيیل CoA
په بایوسینتس LCD کې عکس العمل. لومړی عکس العمل
د اسټیل - CoA ته ترکیب LCD بدلول
ملونيیل CoA. کاتولیک انزایم
دا عکس العمل (اسیلیل کوآ کاربوآکسیلایز) ،
د لیګیسس ټولګي پورې اړه لري. هغه لري
بایلین بندیز. په لومړي کې
د Co2 کوالیټل عکس العمل مرحلې
د انرژی له امله بایوټین سره تړلی
ATP ، په مرحله 2 COO کې لیږدول شوی
په اسیلیل CoA باندې د ملونيیل CoA رامینځته کولو لپاره.
د اسټیل CoA کاربوکسیلاز انزیم فعالیت
د ټولو تعقیب سرعت ټاکي
د ترکیب تعاملات LC
سایټریټ په سیټوسول کې انزیم فعالوي
اسټیل CoA کاربوکسلایز. ملونيیل CoA په
په بدل کې د لوړ لیږد مخه نیسي
غوړ اسیدونه له سیتوسول څخه تر میټریک پورې
مایټوکونډریا د فعالیت مخنیوی کوي
بهرني اسیتیل CoA: کارنیټین اکیلټرانسیفراسیز ،
پدې توګه د لوړ آکسیدریشن بندول
غوړ اسیدونه.
Acetyl-CoA Oxaloacetate →
د HS-CoA سایټریټ

د HSCOA ATP سایټریټ → اسټیل- CoA ADP پای آکسالواسیټیټ
Acetyl-CoA
په سایتوپلازم کې د پیل شوي سبسټراټ په توګه کار کوي
د lcd ترکیب لپاره ، او په اکسلوسیسیټ کې
سایتوسول په کې د بدلونونو څخه تیریږي
د کومې پایلې چې pyruvate رامینځته کیږي.
د کولیسترول بایوسینتیسس
د کولیسترول بایوسینتیسس په اندوپلازمیک ریټیکولم کې واقع کیږي. په مالیکول کې د ټولو کاربن اتومونو سرچینه اسیلیل - سکوآ ده ، کوم چې دلته د مایټوکونډریا څخه د سایټریټ د یوې برخې په توګه راځي ، لکه څنګه چې د غوړ اسیدونو ترکیب کې. د کولیسترول بایوسینتیسس 18 د ATP مالیکولونه او 13 NADPH مالیکولونه مصرفوي.
د کولیسټرول جوړښت له 30 څخه ډیر عکس العمل کې پیښیږي ، کوم چې په څو مرحلو کې ډله کیدی شي.
1. میالوونیک اسید ترکیب.
لومړی دوه ترکیب تعاملات د کیټوجنیز تعاملاتو سره مل دي ، مګر د 3-هایدروکسای-3-میتیلګلوټریل - سکوآ تر ترکیب وروسته انزایم داخلیږي هایډروکسیمیتیل - ګلوټریل - سکو اي رډوکټیس (HMG-SCOA رډوکټیس) ، میالوونیک اسید رامینځته کوي.
 |